144 Superphylum Lophotrochozoa: Flatworms, Rotifers, and Nemerteans
Learning Objectives
By the end of this section, you will be able to do the following:
- Describe the unique anatomical and morphological features of flatworms, rotifers, and Nemertea
- Identify an important extracoelomic cavity found in Nemertea
- Explain the key features of Platyhelminthes and their importance as parasites
Animals belonging to superphylum Lophotrochozoa are triploblastic (have three germ layers) and unlike the cnidarians, they possess an embryonic mesoderm sandwiched between the ectoderm and endoderm. These phyla are also bilaterally symmetrical, meaning that a longitudinal section will divide them into right and left sides that are superficially symmetrical. In these phyla, we also see the beginning of cephalization, the evolution of a concentration of nervous tissues and sensory organs in the head of the organism—exactly where a mobile bilaterally symmetrical organism first encounters its environment.
Lophotrochozoa are also protostomes, in which the blastopore, or the point of invagination of the ectoderm (outer germ layer), becomes the mouth opening into the alimentary canal. This developmental pattern is called protostomy or “first mouth.” Protostomes include acoelomate, pseudocoelomate, and eucoelomate phyla. The coelom is a cavity that separates the ectoderm from the endoderm. In acoelomates, a solid mass of mesoderm is sandwiched between the ectoderm and endoderm and does not form a cavity or “coelom,” leaving little room for organ development; in pseudocoelomates, there is a cavity or pseudocoelom that replaces the blastocoel (the cavity within the blastula), but it is only lined by mesoderm on the outside of the cavity, leaving the gut tube and organs unlined; in eucoelomates, the cavity that obliterates the blastocoel as the coelom develops is lined both on the outside of the cavity (parietal layer) and also on the inside of the cavity, around the gut tube and the internal organs (visceral layer).
Eucoelmate protostomes are schizocoels, in which mesoderm-producing cells typically migrate into the blastocoel during gastrulation and multiply to form a solid mass of cells. Cavities then develop within the cell mass to form the coelom. Since the forming body cavity splits the mesoderm, this protostomic coelom is termed a schizocoelom. As we will see later in this chapter, chordates, the phylum to which we belong, generally develop a coelom by enterocoely: pouches of mesoderm pinch off the invaginating primitive gut, or archenteron, and then fuse to form a complete coelom. We should note here that a eucoelomate can form its “true coelom” by either schizocoely or enterocoely. The process that produces the coelom is different and of taxonomic importance, but the result is the same: a complete, mesodermally lined coelom.
Most organisms placed in the superphylum Lophotrochozoa possess either a lophophore feeding apparatus or a trochophore larvae (thus the contracted name, “lopho-trocho-zoa”). The lophophore is a feeding structure composed of a set of ciliated tentacles surrounding the mouth. A trochophore is a free-swimming larva characterized by two bands of cilia surrounding a top-like body. Some of the phyla classified as Lophotrochozoa may be missing one or both of these defining structures. Nevertheless their placement with the Lophotrochozoa is upheld when ribosomal RNA and other gene sequences are compared. The systematics of this complex group is still unclear and much more work remains to resolve the cladistic relationships among them.
Phylum Platyhelminthes
The flatworms are acoelomate organisms that include many free-living and parasitic forms. The flatworms possess neither a lophophore nor trochophore larvae, although the larvae of one group of flatworms, the Polycladida (named after its many-branched digestive tract), are considered to be homologous to trochophore larvae. Spiral cleavage is also seen in the polycladids and other basal flatworm groups. The developmental pattern of some of the free-living forms is obscured by a phenomenon called “blastomere anarchy,” in which a sort of temporary feeding larva forms, followed by a regrouping of cells within the embryo that gives rise to a second-stage embryo. However, both the monophyly of the flatworms and their placement in the Lophotrochozoa has been supported by molecular analyses.
The Platyhelminthes consist of two monophyletic lineages: the Catenulida and the Rhabditophora. The Catenulida, or “chain worms,” is a small clade of just over 100 species. These worms typically reproduce asexually by budding. However, the offspring do not fully detach from the parents and the formation resembles a chain in appearance. All of the flatworms discussed here are part of the Rhabditophora (“rhabdite bearers”). Rhabdites are rodlike structures discharged in the mucus produced by some free-living flatworms; Eucoelmate protostomes are schizocoels, in which mesoderm-producing cells typically migrate into the blastocoel during gastrulation likely serve in both defense and to provide traction for ciliary gliding along the substrate. Unlike free-living flatworms, many species of trematodes and cestodes are parasitic, including important parasites of humans.

Flatworms have three embryonic tissue layers that give rise to epidermal tissues (from ectoderm), the lining of the digestive system (from endoderm), and other internal tissues (from mesoderm). The epidermal tissue is a single layer of cells or a layer of fused cells (syncytium) that covers two layers of muscle, one circular and the other longitudinal. The mesodermal tissues include mesenchymal cells that contain collagen and support secretory cells that produce mucus and other materials at the surface. Because flatworms are acoelomates, the mesodermal layer forms a solid mass between the outer epidermal surface and the cavity of the digestive system.
Physiological Processes of Flatworms
The free-living species of flatworms are predators or scavengers. Parasitic forms feed by absorbing nutrients provided by their hosts. Most flatworms, such as the planarian shown in (Figure), have a branching gastrovascular cavity rather than a complete digestive system. In such animals, the “mouth” is also used to expel waste materials from the digestive system, and thus also serves as an anus. (A few species may have a second anal pore or opening.) The gut may be a simple sac or highly branched. Digestion is primarily extracellular, with digested materials taken into the cells of the gut lining by phagocytosis. One parasitic group, the tapeworms (cestodes), lacks a digestive system altogether, and absorb digested food from the host.
Flatworms have an excretory system with a network of tubules attached to flame cells, whose cilia beat to direct waste fluids concentrated in the tubules out of the body through excretory pores. The system is responsible for the regulation of dissolved salts and the excretion of nitrogenous wastes. The nervous system consists of a pair of lateral nerve cords running the length of the body with transverse connections between them. Two large cerebral ganglia—concentrations of nerve cell bodies at the anterior end of the worm—are associated with photosensory and chemosensory cells. There is neither a circulatory nor a respiratory system, with gas and nutrient exchange dependent on diffusion and cell-to-cell junctions. This necessarily limits the thickness of the body in these organisms, constraining them to be “flat” worms. Most flatworm species are monoecious (both male and female reproductive organs are found in the same individual), and fertilization is typically internal. Asexual reproduction by fission is common in some groups.
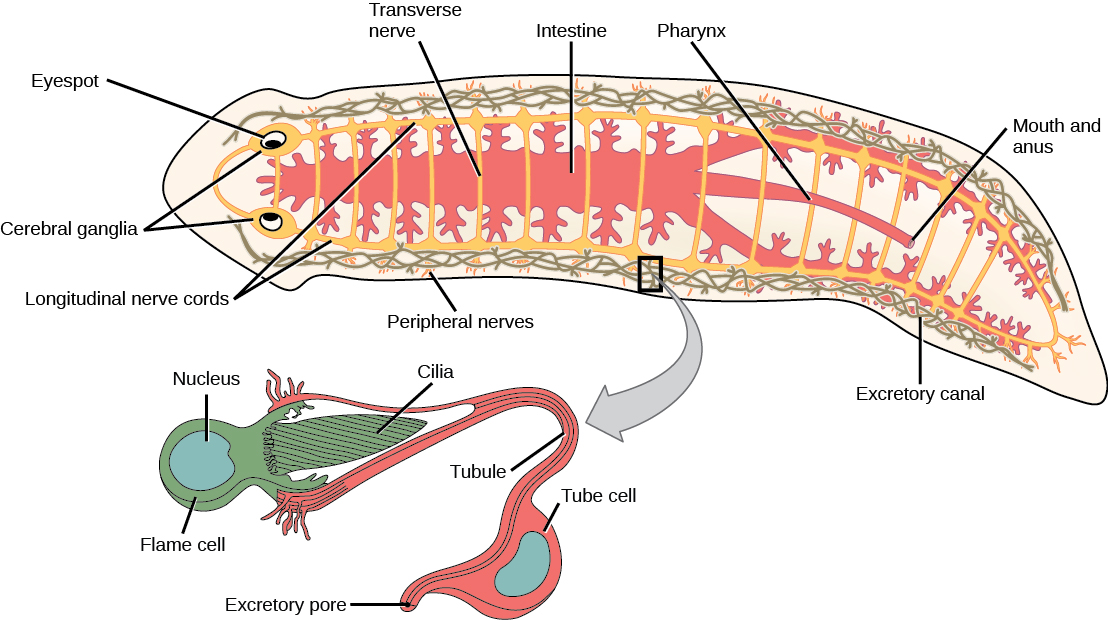
Diversity of Flatworms
The flatworms have been traditionally divided into four classes: Turbellaria, Monogenea, Trematoda, and Cestoda ((Figure)). However, the relationships among members of these classes has recently been reassessed, with the turbellarians in particular now viewed as paraphyletic, since its descendants may also include members of the other three classes. Members of the clade or class Rhabditophora are now dispersed among multiple orders of Platyhelminthes, the most familiar of these being the Polycladida, which contains the large marine flatworms; the Tricladida (which includes Dugesia [“planaria”] and Planaria and its relatives); and the major parasitic orders: Monogenea (fish ectoparasites), Trematoda (flukes), and Cestoda (tapeworms), which together form a monophyletic clade.
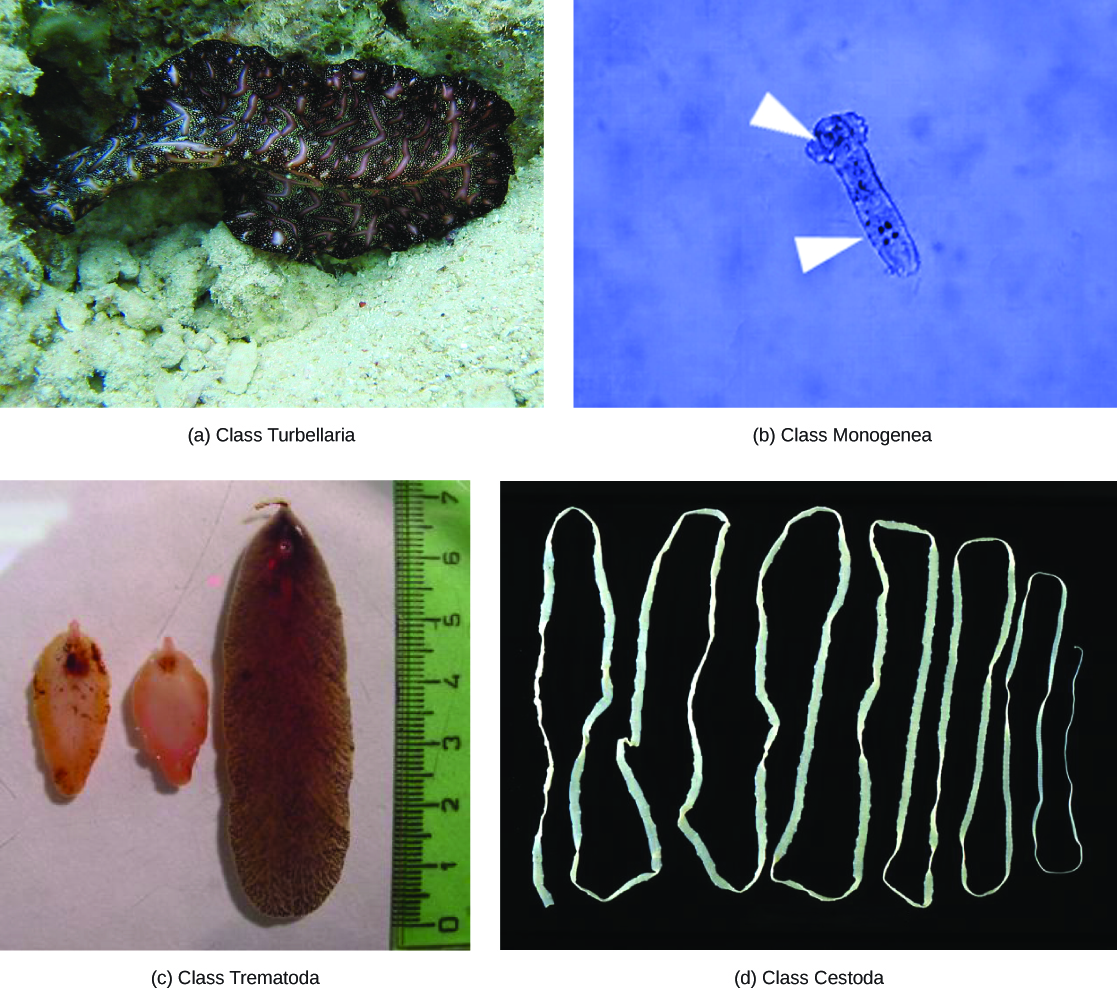
Most free-living flatworms are marine polycladids, although tricladid species live in freshwater or moist terrestrial environments, and there are a number of members from other orders in both environments. The ventral epidermis of free-living flatworms is ciliated, which facilitates their locomotion. Some free-living flatworms are capable of remarkable feats of regeneration in which an individual may regrow its head or tail after being severed, or even several heads if the planaria is cut lengthwise.
The monogeneans are ectoparasites, mostly of fish, with simple life cycles that consist of a free-swimming larva that attaches to a fish, prior to its transformation to the ectoparasitic adult form. The parasite has only one host and that host is usually very specific. The worms may produce enzymes that digest the host tissues, or they may simply graze on surface mucus and skin particles. Most monogeneans are hermaphroditic, but the male gametes develop first and so cross-fertilization is quite common.
The trematodes, or flukes, are internal parasites of mollusks and many other groups, including humans. Trematodes have complex life cycles that involve a primary host in which sexual reproduction occurs, and one or more secondary hosts in which asexual reproduction occurs. The primary host is usually a vertebrate and the secondary host is almost always a mollusk, in which multiple larvae are produced asexually. Trematodes, which attached internally to the host via an oral and medial sucker, are responsible for serious human diseases including schistosomiasis, caused by several species of the blood fluke, Schistosoma spp. Various forms of schistosomiasis infect an estimated 200 million people in the tropics, leading to organ damage, secondary infection by bacteria, and chronic symptoms like fatigue. Infection occurs when the human enters the water and metacercaria larvae, released from the snail host, locate and penetrate the skin. The parasite infects various organs in the body and feeds on red blood cells before reproducing.
Many of the eggs are released in feces and find their way into a waterway, where they are able to reinfect the snail host. The eggs, which have a barb on them, can damage the vascular system of the human host, causing ulceration, abscesses, and bloody diarrhea, wherever they reside, thereby allowing other pathogens to cause secondary infections. In fact, it is the parasite’s eggs that produce most of the main ill effects of schistosomiasis. Many eggs do not make the transit through the veins of the host for elimination, and are swept by blood flow back to the liver and other locations, where they can cause severe inflammation. In the liver, the errant eggs may impede circulation and cause cirrhosis. Control is difficult in impoverished areas in unsanitary, crowded conditions, and prognosis is poor in people with heavy infections of Schistosoma japonicum, without early treatment.
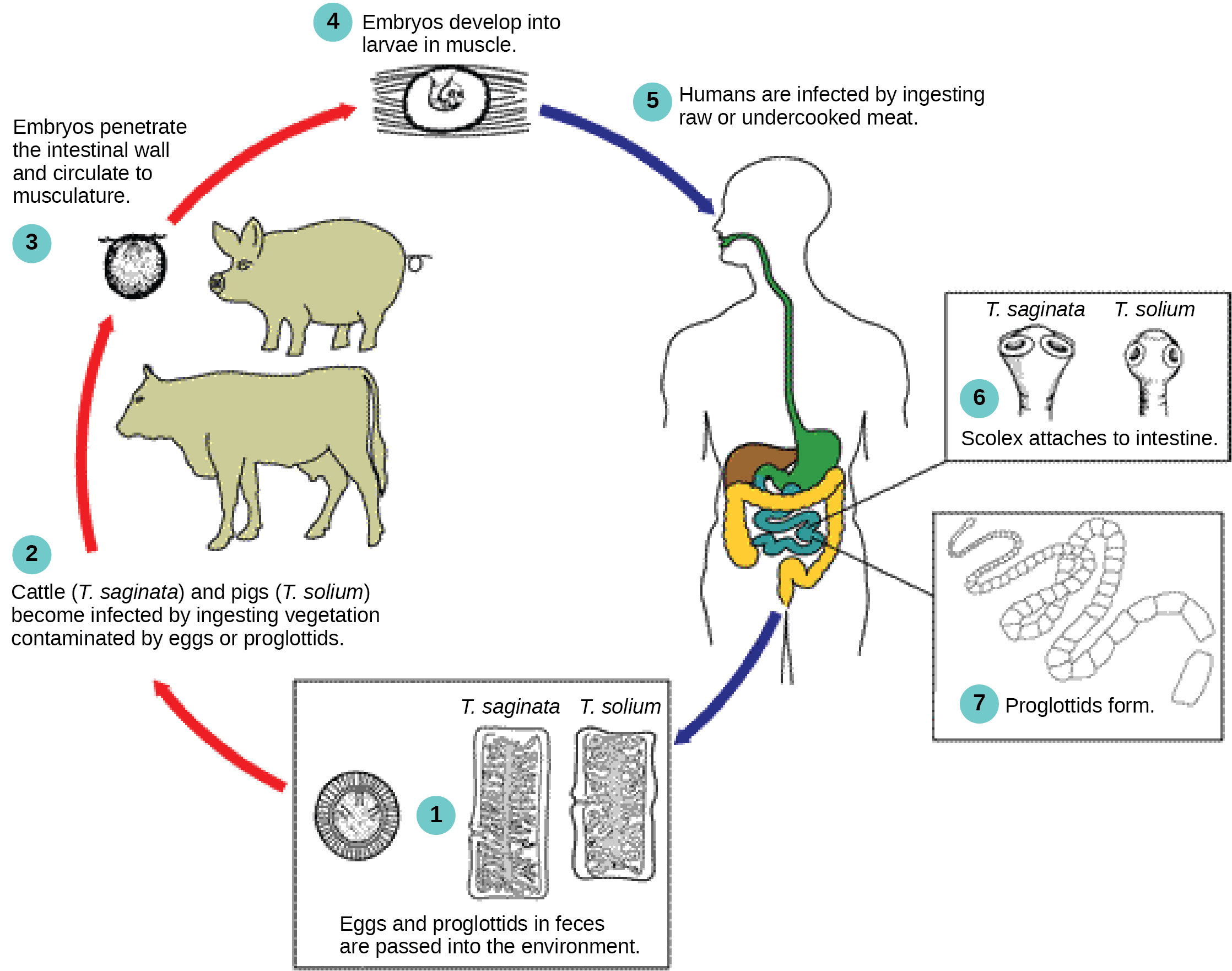
The cestodes, or tapeworms, are also internal parasites, mainly of vertebrates ((Figure)). Tapeworms, such as those of Taenia spp, live in the intestinal tract of the primary host and remain fixed using a sucker or hooks on the anterior end, or scolex, of the tapeworm body, which is essentially a colony of similar subunits called proglottids. Each proglottid may contain an excretory system with flame cells, along with reproductive structures, both male and female. Because they are so long and flat, tapeworms do not need a digestive system; instead, they absorb nutrients from the food matter surrounding them in the host’s intestine by diffusion.
Proglottids are produced at the scolex and gradually migrate to the end of the tapeworm; at this point, they are “mature” and all structures except fertilized eggs have degenerated. Most reproduction occurs by cross-fertilization between different worms in the same host, but may also occur between proglottids. The mature proglottids detach from the body of the worm and are released into the feces of the organism. The eggs are eaten by an intermediate host, typically another vertebrate. The juvenile worm infects the intermediate host and takes up residence, usually in muscle tissue. When the muscle tissue is consumed by the primary host, the cycle is completed. There are several tapeworm parasites of humans that are transmitted by eating uncooked or poorly cooked pork, beef, or fish.
Phylum Rotifera
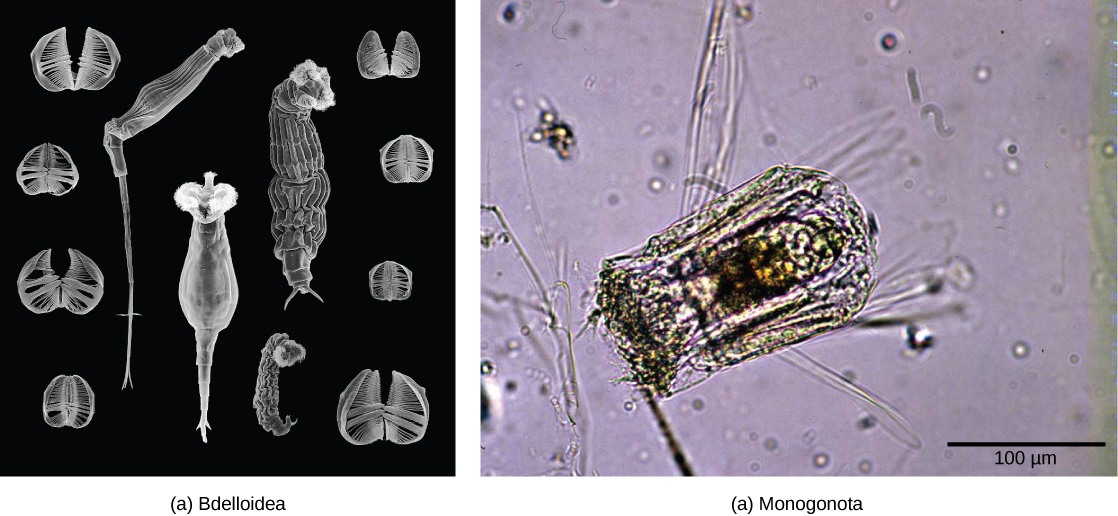
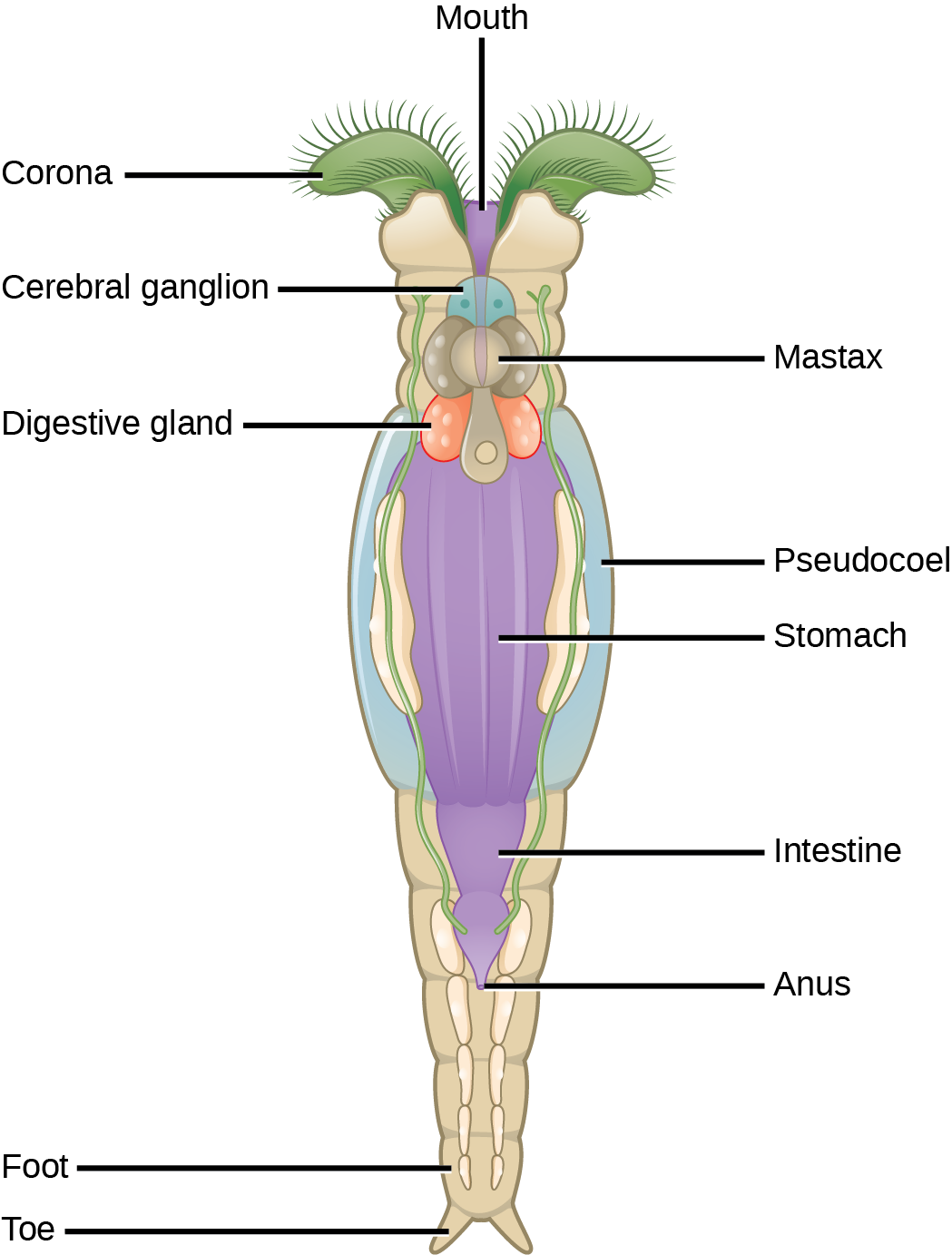
The rotifers (“wheel-bearer”) belong to a group of microscopic (about 100 µm to 2 mm) mostly aquatic animals that get their name from the corona—a pair of ciliated feeding structures that appear to rotate when viewed under the light microscope ((Figure)). Although their taxonomic status is currently in flux, one treatment places the rotifers in three classes: Bdelloidea, Monogononta, and Seisonidea. In addition, the parasitic “spiny headed worms” currently in phylum Acanthocephala, appear to be modified rotifers and will probably be placed into the group in the near future. Undoubtedly the rotifers will continue to be revised as more phylogenetic evidence becomes available.
The pseudocoelomate body of a rotifer is remarkably complex for such a small animal (roughly the size of a Paramecium) and is divided into three sections: a head (which contains the corona), a trunk (which contains most of the internal organs), and the foot. A cuticle, rigid in some species and flexible in others, covers the body surface. They have both skeletal muscle associated with locomotion and visceral muscles associated with the gut, both composed of single cells. Rotifers are typically free-swimming or planktonic (drifting) organisms, but the toes or extensions of the foot can secrete a sticky material to help them adhere to surfaces. The head contains a number of eyespots and a bilobed “brain,” with nerves extending into the body.
Rotifers are commonly found in freshwater and some saltwater environments throughout the world. As filter feeders, they will eat dead material, algae, and other microscopic living organisms, and are therefore very important components of aquatic food webs. A rotifer’s food is directed toward the mouth by the current created from the movement of the coronal cilia. The food particles enter the mouth and travel first to the mastax—a muscular pharynx with toothy jaw-like structures. Examples of the jaws of various rotifers are seen in (Figure)a. Masticated food passes near digestive and salivary glands, into the stomach, and then to the intestines. Digestive and excretory wastes are collected in a cloacal bladder before being released out the anus.
Watch this video to see rotifers feeding.
About 2,200 species of rotifers have been identified. (Figure) shows the anatomy of a rotifer belonging to class Bdelloidea. Some rotifers are dioecious organisms and exhibit sexual dimorphism (males and females have different forms). In many dioecious species, males are short-lived and smaller with no digestive system and a single testis. Many rotifer species exhibit haplodiploidy, a method of sex determination in which a fertilized egg develops into a female and an unfertilized egg develops into a male. However, reproduction in the bdelloid rotifers is exclusively parthenogenetic and appears to have been so for millions of years: Thus, all bdelloid rotifers and their progeny are female! The bdelloids may compensate for this genetic insularity by borrowing genes from the DNA of other species. Up to 10% of a bdelloid genome comprises genes imported from related species. Some rotifer eggs are capable of extended dormancy for protection during harsh environmental conditions.
Phylum Nemertea
The Nemertea are colloquially known as ribbon worms or proboscis worms. Most species of phylum Nemertea are marine and predominantly benthic (bottom dwellers), with an estimated 900 known species. However, nemerteans have been recorded in freshwater and very damp terrestrial habitats as well. Most nemerteans are carnivores, feeding on worms, clams, and crustaceans. Some species are scavengers, and some, like Malacobdella grossa, have also evolved commensal relationships with mollusks. Economically important species have at times devastated commercial fishing of clams and crabs. Nemerteans have almost no predators and two species are sold as fish bait.
Morphology
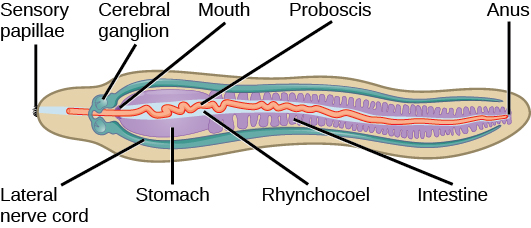
Nemerteans vary in size from 1 cm to several meters. They show bilateral symmetry and remarkable contractile properties. Because of their contractility, they can change their morphological presentation in response to environmental cues. Animals in phylum Nemertea are soft and unsegmented animals, with a morphology like a flattened tube. ((Figure)).

A unique characteristic of this phylum is the presence of an eversible proboscis enclosed in a pocket called a rhynchocoel (not part of the animal’s actual coelom). The proboscis is located dorsal to the gut and serves as a harpoon or tentacle for food capture. In some species it is ornamented with barbs. The rhynchocoel is a fluid-filled cavity that extends from the head to nearly two-thirds of the length of the gut in these animals ((Figure)). The proboscis may be extended by hydrostatic pressure generated by contraction of muscle of the rhynchocoel and retracted by a retractor muscle attached to the rear wall of the rhynchocoel.
Watch this video to see a nemertean attack a polychaete with its proboscis.
Digestive System
The nemerteans, which are primarily predators of annelids and crustaceans, have a well-developed digestive system. A mouth opening that is ventral to the rhynchocoel leads into the foregut, followed by the intestine. The intestine is present in the form of diverticular pouches and ends in a rectum that opens via an anus. Gonads are interspersed with the intestinal diverticular pouches and open outward via genital pores.
Nemerteans are sometimes classified as acoels, but because their closed circulatory system is derived from the coelomic cavity of the embryo, they may be regarded as coelomic. Their circulatory system consists of a closed loop formed by a connected pair of lateral blood vessels. Some species may also have a dorsal vessel or cross-connecting vessels in addition to lateral ones. Although the circulatory fluid contains cells, it is often colorless. However, the blood cells of some species bear hemoglobin as well as other yellow or green pigments. The blood vessels are contractile, although there is usually no regular circulatory pathway, and movement of blood is also facilitated by the contraction of muscles in the body wall. The circulation of fluids in the rhychocoel is more or less independent of the blood circulation, although blind branches from the blood vessels into the rhyncocoel wall can mediate exchange of materials between them. A pair of protonephridia, or excretory tubules, is present in these animals to facilitate osmoregulation. Gaseous exchange occurs through the skin.
Nervous System
Nemerteans have a “brain” composed of four ganglia situated at the anterior end, around the rhynchocoel. Paired longitudinal nerve cords emerge from the brain ganglia and extend to the posterior end. Additional nerve cords are found in some species. Interestingly, the brain can contain hemoglobin, which acts as an oxygen reserve. Ocelli or eyespots are present in pairs, in multiples of two in the anterior portion of the body. It is speculated that the eyespots originate from neural tissue and not from the epidermis.
Reproduction
Nemerteans, like flatworms, have excellent powers of regeneration, and asexual reproduction by fragmentation is seen in some species. Most animals in phylum Nemertea are dioecious, although freshwater species may be hermaphroditic. Stem cells that become gametes aggregate within gonads placed along the digestive tract. Eggs and sperm are released into the water, and fertilization occurs externally. Like most lophotrochozoan protostomes, cleavage is spiral, and development is usually direct, although some species have a trochophore-like larva, in which a young worm is constructed from a series of imaginal discs that begin as invaginations from the body surface of the larva.
Section Summary
This section describes three phyla of relatively simple invertebrates: one acoelomate, one pseudocoelomate, and one eucoelomate. Flatworms are acoelomate, triploblastic animals. They lack circulatory and respiratory systems, and have a rudimentary excretory system. This digestive system is incomplete in most species, and absent in tapeworms. There are four traditional groups of flatworms, the largely free-living turbellarians, which include polycladid marine worms and tricladid freshwater species, the ectoparasitic monogeneans, and the endoparasitic trematodes and cestodes. Trematodes have complex life cycles involving a molluscan secondary host and a primary host in which sexual reproduction takes place. Cestodes, or tapeworms, infect the digestive systems of their primary vertebrate hosts.
Rotifers are microscopic, multicellular, mostly aquatic organisms that are currently under taxonomic revision. The group is characterized by the ciliated, wheel-like corona, located on their head. Food collected by the corona is passed to another structure unique to this group of organisms—the mastax or jawed pharynx.
The nemerteans are probably simple eucoelomates. These ribbon-shaped animals also bear a specialized proboscis enclosed within a rhynchocoel. The development of a closed circulatory system derived from the coelom is a significant difference seen in this species compared to other phyla described here. Alimentary, nervous, and excretory systems are more developed in the nemerteans than in the flatworms or rotifers. Embryonic development of nemertean worms proceeds via a planuliform or trochophore-like larval stage.
Review Questions
Which group of flatworms are primarily ectoparasites of fish?
- monogeneans
- trematodes
- cestodes
- turbellarians
A
The rhynchocoel is a ________.
- circulatory system
- fluid-filled cavity
- primitive excretory system
- proboscis
B
Critical Thinking Questions
How does the tapeworm body plan support widespread dissemination of the parasite?
The tapeworm bodies are made of a series of small subunits called proglottids. Each proglottid contains a sexually mature reproductive system that can cross-fertilize within a single tapeworm. Proglottids containing fertilized eggs break off the tapeworm, and are transported to the environment to find a new host. Thus, each segment of a tapeworm’s body is capable of infecting new hosts.
Glossary
- corona
- wheel-like structure on the anterior portion of the rotifer that contains cilia and moves food and water toward the mouth
- mastax
- jawed pharynx unique to the rotifers
- Nemertea
- phylum of dorsoventrally flattened protostomes known as ribbon worms
- parapodium
- fleshy, flat, appendage that protrudes in pairs from each segment of polychaetes
- pilidium
- larval form found in some nemertine species
- planuliform
- larval form found in phylum Nemertea
- rhynchocoel
- cavity present above the mouth that houses the proboscis
- schizocoelom
- coelom formed by groups of cells that split from the endodermal layer
