138 Features Used to Classify Animals
Learning Objectives
By the end of this section, you will be able to do the following:
- Explain the differences in animal body plans that support basic animal classification
- Compare and contrast the embryonic development of protostomes and deuterostomes
<!–<anchor id=”ch27_module_2″/>–>Scientists have developed a classification scheme that categorizes all members of the animal kingdom, although there are exceptions to most “rules” governing animal classification ((Figure)). Animals have been traditionally classified according to two characteristics: body plan and developmental pathway. The major feature of the body plan is its symmetry: how the body parts are distributed along the major body axis. Symmetrical animals can be divided into roughly equivalent halves along at least one axis. Developmental characteristics include the number of germ tissue layers formed during development, the origin of the mouth and anus, the presence or absence of an internal body cavity, and other features of embryological development, such as larval types or whether or not periods of growth are interspersed with molting.
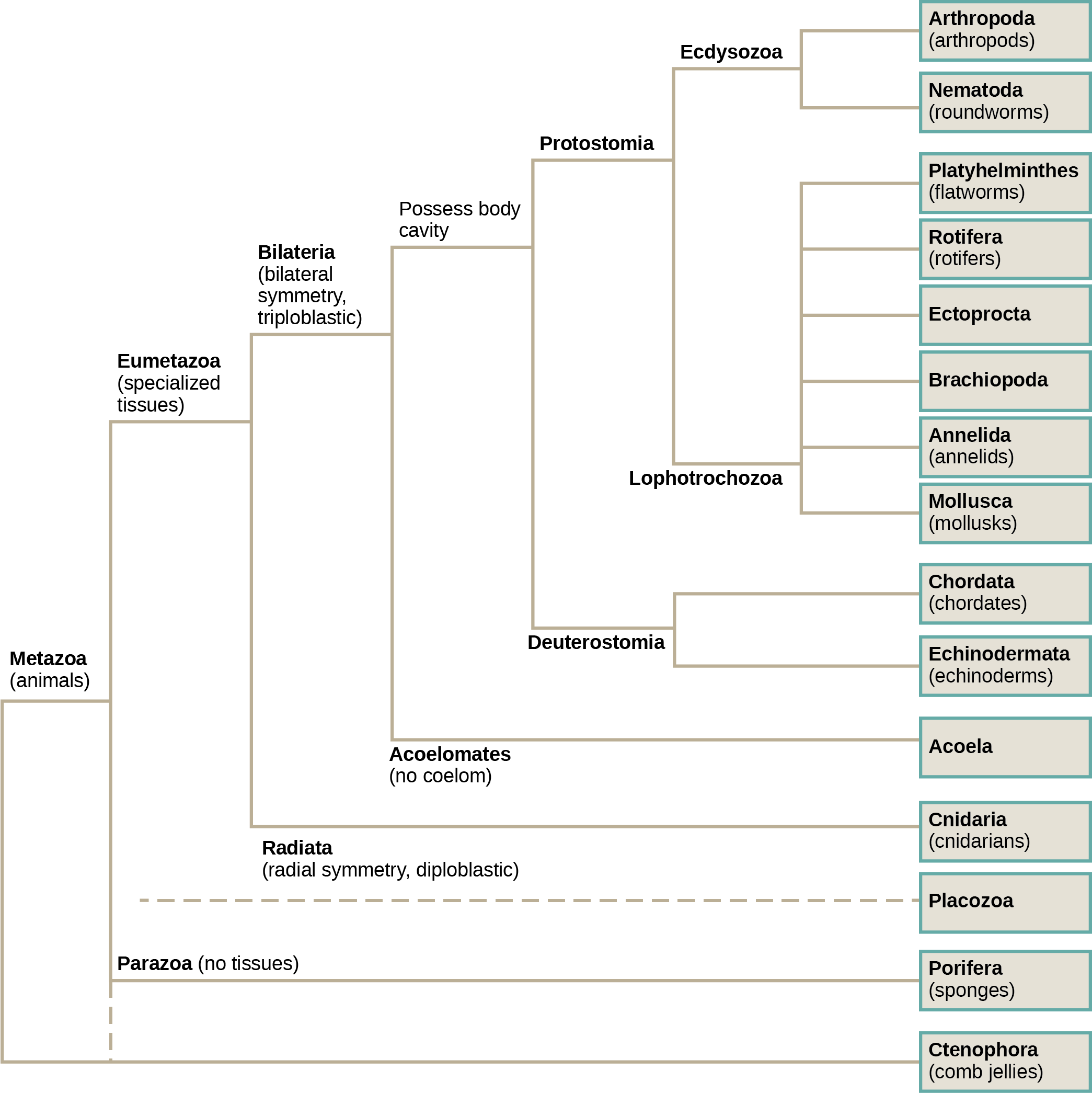
Which of the following statements is false?
- Eumetazoans have specialized tissues and parazoans don’t.
- Lophotrochozoa and Ecdysozoa are both Bilataria.
- Acoela and Cnidaria both possess radial symmetry.
- Arthropods are more closely related to nematodes than they are to annelids.
<!–<para>C–>
Animal Characterization Based on Body Symmetry
At a very basic level of classification, true animals can be largely divided into three groups based on the type of symmetry of their body plan: radially symmetrical, bilaterally symmetrical, and asymmetrical. Asymmetry is seen in two modern clades, the Parazoa ((Figure)a) and Placozoa. (Although we should note that the ancestral fossils of the Parazoa apparently exhibited bilateral symmetry.) One clade, the Cnidaria ((Figure)b,c), exhibits radial or biradial symmetry: Ctenophores have rotational symmetry ((Figure)e). Bilateral symmetry is seen in the largest of the clades, the Bilateria ((Figure)d); however the Echinodermata are bilateral as larvae and metamorphose secondarily into radial adults. All types of symmetry are well suited to meet the unique demands of a particular animal’s lifestyle.
Radial symmetry is the arrangement of body parts around a central axis, as is seen in a bicycle wheel or pie. It results in animals having top and bottom surfaces but no left and right sides, nor front or back. If a radially symmetrical animal is divided in any direction along the oral/aboral axis (the side with a mouth is “oral side,” and the side without a mouth is the “aboral side”), the two halves will be mirror images. This form of symmetry marks the body plans of many animals in the phyla Cnidaria, including jellyfish and adult sea anemones ((Figure)b, c). Radial symmetry equips these sea creatures (which may be sedentary or only capable of slow movement or floating) to experience the environment equally from all directions. Bilaterally symmetrical animals, like butterflies ((Figure)d) have only a single plane along which the body can be divided into equivalent halves. The Ctenophora ((Figure)e), although they look similar to jellyfish, are considered to have rotational symmetry rather than radial or biradial symmetry because division of the body into two halves along the oral/aboral axis divides them into two copies of the same half, with one copy rotated 180o, rather than two mirror images.

Bilateral symmetry involves the division of the animal through a midsagittal plane, resulting in two superficially mirror images, right and left halves, such as those of a butterfly ((Figure)d), crab, or human body. Animals with bilateral symmetry have a “head” and “tail” (anterior vs. posterior), front and back (dorsal vs. ventral), and right and left sides ((Figure)). All Eumetazoa except those with secondary radial symmetry are bilaterally symmetrical. The evolution of bilateral symmetry that allowed for the formation of anterior and posterior (head and tail) ends promoted a phenomenon called cephalization, which refers to the collection of an organized nervous system at the animal’s anterior end. In contrast to radial symmetry, which is best suited for stationary or limited-motion lifestyles, bilateral symmetry allows for streamlined and directional motion. In evolutionary terms, this simple form of symmetry promoted active and controlled directional mobility and increased sophistication of resource-seeking and predator-prey relationships.
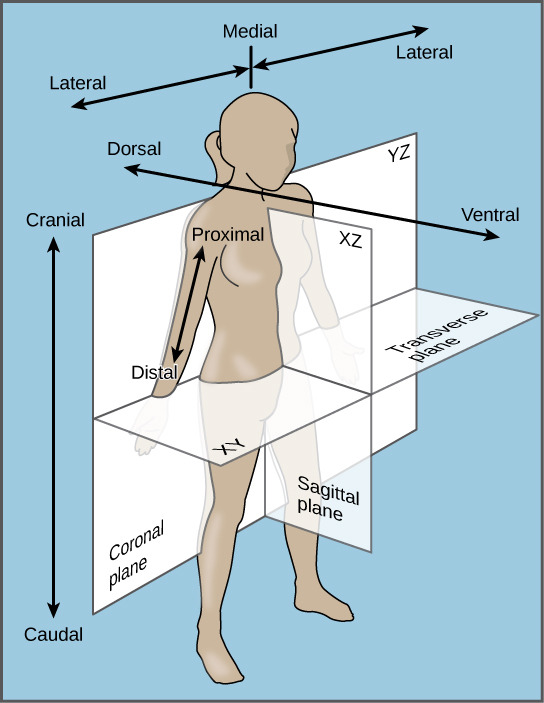
Animals in the phylum Echinodermata (such as sea stars, sand dollars, and sea urchins) display modified radial symmetry as adults, but as we have noted, their larval stages (such as the bipinnaria) initially exhibit bilateral symmetry until they metamorphose in animals with radial symmetry (this is termed secondary radial symmetry). Echinoderms evolved from bilaterally symmetrical animals; thus, they are classified as bilaterally symmetrical.
Watch this video to see a quick sketch of the different types of body symmetry.
Animal Characterization Based on Features of Embryological Development
Most animal species undergo a separation of tissues into germ layers during embryonic development. Recall that these germ layers are formed during gastrulation, and that each germ layer typically gives rise to specific types of embryonic tissues and organs. Animals develop either two or three embryonic germ layers ((Figure)). The animals that display radial, biradial, or rotational symmetry develop two germ layers, an inner layer (endoderm or mesendoderm) and an outer layer (ectoderm). These animals are called diploblasts, and have a nonliving middle layer between the endoderm and ectoderm (although individual cells may be distributed through this middle layer, there is no coherent third layer of tissue). The four clades considered to be diploblastic have different levels of complexity and different developmental pathways, although there is little information about development in Placozoa. More complex animals (usually those with bilateral symmetry) develop three tissue layers: an inner layer (endoderm), an outer layer (ectoderm), and a middle layer (mesoderm). Animals with three tissue layers are called triploblasts.
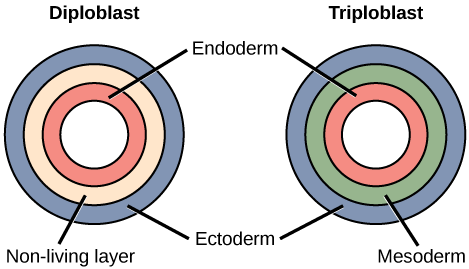
Which of the following statements about diploblasts and triploblasts is false?
- Animals that display only radial symmetry during their lifespans are diploblasts.
- Animals that display bilateral symmetry are triploblasts.
- The endoderm gives rise to the lining of the digestive tract and the respiratory tract.
- The mesoderm gives rise to the central nervous system.
<!–<para> D–>
Each of the three germ layers is programmed to give rise to specific body tissues and organs, although there are variations on these themes. Generally speaking, the endoderm gives rise to the lining of the digestive tract (including the stomach, intestines, liver, and pancreas), as well as to the lining of the trachea, bronchi, and lungs of the respiratory tract, along with a few other structures. The ectoderm develops into the outer epithelial covering of the body surface, the central nervous system, and a few other structures. The mesoderm is the third germ layer; it forms between the endoderm and ectoderm in triploblasts. This germ layer gives rise to all specialized muscle tissues (including the cardiac tissues and muscles of the intestines), connective tissues such as the skeleton and blood cells, and most other visceral organs such as the kidneys and the spleen. Diploblastic animals may have cell types that serve multiple functions, such as epitheliomuscular cells, which serve as a covering as well as contractile cells.
Presence or Absence of a Coelom
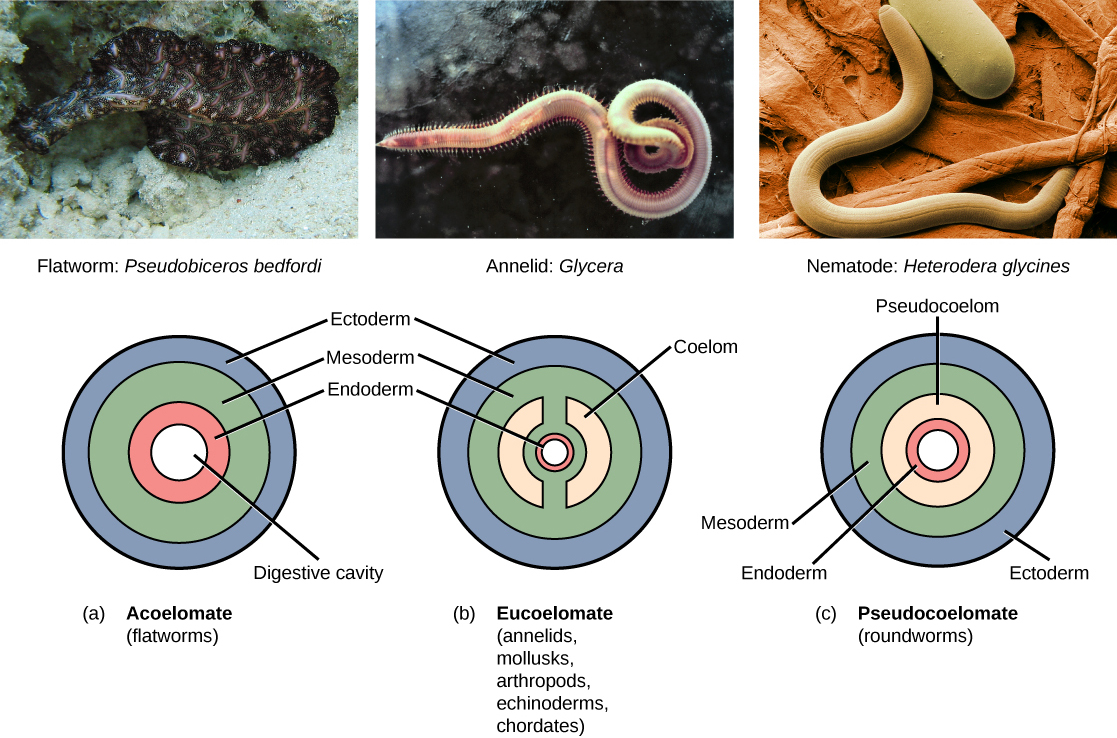
Further subdivision of animals with three germ layers (triploblasts) results in the separation of animals that may develop an internal body cavity derived from mesoderm, called a coelom, and those that do not. This epithelial cell-lined coelomic cavity, usually filled with fluid, lies between the visceral organs and the body wall. It houses many organs such as the digestive, urinary, and reproductive systems, the heart and lungs, and also contains the major arteries and veins of the circulatory system. In mammals, the body cavity is divided into the thoracic cavity, which houses the heart and lungs, and the abdominal cavity, which houses the digestive organs. In the thoracic cavity further subdivision produces the pleural cavity, which provides space for the lungs to expand during breathing, and the pericardial cavity, which provides room for movements of the heart. The evolution of the coelom is associated with many functional advantages. For example, the coelom provides cushioning and shock absorption for the major organ systems that it encloses. In addition, organs housed within the coelom can grow and move freely, which promotes optimal organ development and placement. The coelom also provides space for the diffusion of gases and nutrients, as well as body flexibility, promoting improved animal motility.
Triploblasts that do not develop a coelom are called acoelomates, and their mesoderm region is completely filled with tissue, although they do still have a gut cavity. Examples of acoelomates include animals in the phylum Platyhelminthes, also known as flatworms. Animals with a true coelom are called eucoelomates (or coelomates) ((Figure)). In such cases, a true coelom arises entirely within the mesoderm germ layer and is lined by an epithelial membrane. This membrane also lines the organs within the coelom, connecting and holding them in position while allowing them some freedom of movement. Annelids, mollusks, arthropods, echinoderms, and chordates are all eucoelomates. A third group of triploblasts has a slightly different coelom lined partly by mesoderm and partly by endoderm. Although still functionally a coelom, these are considered “false” coeloms, and so we call these animals pseudocoelomates. The phylum Nematoda (roundworms) is an example of a pseudocoelomate. True coelomates can be further characterized based on other features of their early embryological development.
Embryonic Development of the Mouth
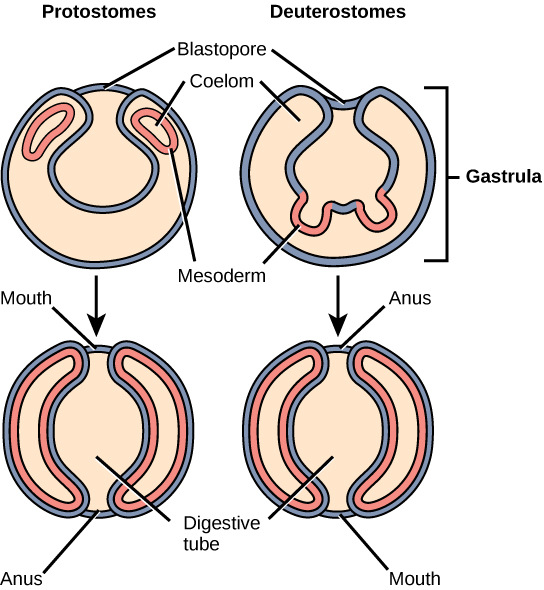
Bilaterally symmetrical, tribloblastic eucoelomates can be further divided into two groups based on differences in the origin of the mouth. When the primitive gut forms, the opening that first connects the gut cavity to the outside of the embryo is called the blastopore. Most animals have openings at both ends of the gut: mouth at one end and anus at the other. One of these openings will develop at or near the site of the blastopore. In Protostomes (“mouth first”), the mouth develops at the blastopore ((Figure)). In Deuterostomes (“mouth second”), the mouth develops at the other end of the gut ((Figure)) and the anus develops at the site of the blastopore. Protostomes include arthropods, mollusks, and annelids. Deuterostomes include more complex animals such as chordates but also some “simple” animals such as echinoderms. Recent evidence has challenged this simple view of the relationship between the location of the blastopore and the formation of the mouth, however, and the theory remains under debate. Nevertheless, these details of mouth and anus formation reflect general differences in the organization of protostome and deuterostome embryos, which are also expressed in other developmental features.
One of these differences between protostomes and deuterostomes is the method of coelom formation, beginning from the gastrula stage. Since body cavity formation tends to accompany the formation of the mesoderm, the mesoderm of protostomes and deuterostomes forms differently. The coelom of most protostomes is formed through a process called schizocoely. The mesoderm in these organisms is usually the product of specific blastomeres, which migrate into the interior of the embryo and form two clumps of mesodermal tissue. Within each clump, cavities develop and merge to form the hollow opening of the coelom. Deuterostomes differ in that their coelom forms through a process called enterocoely. Here, the mesoderm develops as pouches that are pinched off from the endoderm tissue. These pouches eventually fuse and expand to fill the space between the gut and the body wall, giving rise to the coelom.
Another difference in organization of protostome and deuterostome embryos is expressed during cleavage. Protostomes undergo spiral cleavage, meaning that the cells of one pole of the embryo are rotated, and thus misaligned, with respect to the cells of the opposite pole. This is due to the oblique angle of cleavage relative to the two poles of the embryo. Deuterostomes undergo radial cleavage, where the cleavage axes are either parallel or perpendicular to the polar axis, resulting in the parallel (up-and-down) alignment of the cells between the two poles.
A second distinction between the types of cleavage in protostomes and deuterostomes relates to the fate of the resultant blastomeres (cells produced by cleavage). In addition to spiral cleavage, protostomes also undergo determinate cleavage. This means that even at this early stage, the developmental fate of each embryonic cell is already determined. A given cell does not have the ability to develop into any cell type other than its original destination. Removal of a blastomere from an embryo with determinate cleavage can result in missing structures, and embryos that fail to develop. In contrast, deuterostomes undergo indeterminate cleavage, in which cells are not yet fully committed at this early stage to develop into specific cell types. Removal of individual blastomeres from these embryos does not result in the loss of embryonic structures. In fact, twins (clones) can be produced as a result from blastomeres that have been separated from the original mass of blastomere cells. Unlike protostomes, however, if some blastomeres are damaged during embryogenesis, adjacent cells are able to compensate for the missing cells, and the embryo is not damaged. These cells are referred to as undetermined cells. This characteristic of deuterostomes is reflected in the existence of familiar embryonic stem cells, which have the ability to develop into any cell type until their fate is programmed at a later developmental stage.
The Evolution of the CoelomOne of the first steps in the classification of animals is to examine the animal’s body. One structure that is used in classification of animals is the body cavity or coelom. The body cavity develops within the mesoderm, so only triploblastic animals can have body cavities. Therefore body cavities are found only within the Bilateria. In other animal clades, the gut is either close to the body wall or separated from it by a jelly-like material. The body cavity is important for two reasons. Fluid within the body cavity protects the organs from shock and compression. In addition, since in triploblastic embryos, most muscle, connective tissue, and blood vessels develop from mesoderm, these tissues developing within the lining of the body cavity can reinforce the gut and body wall, aid in motility, and efficiently circulate nutrients.
To recap what we have discussed above, animals that do not have a coelom are called acoelomates. The major acoelomate group in the Bilateria is the flatworms, including both free-living and parasitic forms such as tapeworms. In these animals, mesenchyme fills the space between the gut and the body wall. Although two layers of muscle are found just under the epidermis, there is no muscle or other mesodermal tissue around the gut. Flatworms rely on passive diffusion for nutrient transport across their body.
In pseudocoelomates, there is a body cavity between the gut and the body wall, but only the body wall has mesodermal tissue. In these animals, the mesoderm forms, but does not develop cavities within it. Major pseudocoelomate phyla are the rotifers and nematodes. Animals that have a true coelom are called eucoelomates; all vertebrates, as well as molluscs, annelids, arthropods, and echinoderms, are eucoelomates. The coelom develops within the mesoderm during embryogenesis. Of the major bilaterian phyla, the molluscs, annelids, and arthropods are schizocoels, in which the mesoderm splits to form the body cavity, while the echinoderms and chordates are enterocoels, in which the mesoderm forms as two or more buds off of the gut. These buds separate from the gut and coalesce to form the body cavity. In the vertebrates, mammals have a subdivided body cavity, with the thoracic cavity separated from the abdominal cavity. The pseudocoelomates may have had eucoelomate ancestors and may have lost their ability to form a complete coelom through genetic mutations. Thus, this step in early embryogenesis—the formation of the coelom—has had a large evolutionary impact on the various species of the animal kingdom.
Section Summary
Organisms in the animal kingdom are classified based on their body morphology, their developmental pathways, and their genetic affinities. The relationships between the Eumetazoa and more basal clades (Ctenophora, Porifera, and Placozoa) are still being debated. The Eumetazoa (“true animals”) are divided into those with radial versus bilateral symmetry. Generally, the simpler and often nonmotile animals display radial symmetry, which allows them to explore their environment in all directions. Animals with radial symmetry are also generally characterized by the development of two embryological germ layers, the endoderm and ectoderm, whereas animals with bilateral symmetry are generally characterized by the development of a third embryologic germ layer, the mesoderm. Animals with three germ layers, called triploblasts, are further characterized by the presence or absence of an internal body cavity called a coelom. The presence of a coelom affords many advantages, and animals with a coelom may be termed true coelomates or pseudocoelomates, depending the extent to which mesoderm lines the body cavity. Coelomates are further divided into one of two groups called protostomes and deuterostomes, based on a number of developmental characteristics, including differences in zygote cleavage, the method of coelom formation, and the rigidity of the developmental fate of blastomeres.
Visual Connection Questions
(Figure) Which of the following statements is false?
- Eumetazoans have specialized tissues and parazoans don’t.
- Lophotrochozoa and Ecdysozoa are both Bilataria.
- Acoela and Cnidaria both possess radial symmetry.
- Arthropods are more closely related to nematodes than they are to annelids.
(Figure) C
(Figure) Which of the following statements about diploblasts and triploblasts is false?
- Animals that display radial symmetry are diploblasts.
- Animals that display bilateral symmetry are triploblasts.
- The endoderm gives rise to the lining of the digestive tract and the respiratory tract.
- The mesoderm gives rise to the central nervous system.
(Figure) D
Review Questions
Which of the following organisms is most likely to be a diploblast?
- sea star
- shrimp
- jellyfish
- insect
C
Which of the following is not possible?
- radially symmetrical diploblast
- diploblastic eucoelomate
- protostomic coelomate
- bilaterally symmetrical deuterostome
B
An animal whose development is marked by radial cleavage and enterocoely is ________.
- a deuterostome
- an annelid or mollusk
- either an acoelomate or eucoelomate
- none of the above
A
Critical Thinking Questions
Using the following terms, explain what classifications and groups humans fall into, from the most general to the most specific: symmetry, germ layers, coelom, cleavage, embryological development.
Humans have body plans that are bilaterally symmetrical and are characterized by the development of three germ layers, making them triploblasts. Humans have true coeloms and are thus eucoelomates. As deuterostomes, humans are characterized by radial and indeterminate cleavage.
Explain some of the advantages brought about through the evolution of bilateral symmetry and coelom formation.
The evolution of bilateral symmetry led to designated head and tail body regions, and promoted more efficient mobility for animals. This improved mobility allowed for more skillful seeking of resources and prey escaping from predators. The appearance of the coelom in coelomates provides many internal organs with shock absorption, making them less prone to physical damage from bodily assault. A coelom also gives the body greater flexibility, which promotes more efficient movement. The relatively loose placement of organs within the coelom allows them to develop and grow with some spatial freedom, which promoted the evolution of optimal organ arrangement. The coelom also provides space for a circulatory system, which is an advantageous way to distribute body fluids and gases.
Glossary
- acoelomate
- animal without a body cavity
- bilateral symmetry
- type of symmetry in which there is only one plane of symmetry, so the left and right halves of an animal are mirror images
- blastopore
- opening into the archenteron that forms during gastrulation
- coelom
- lined body cavity
- determinate cleavage
- cleavage pattern in which developmental fate of each blastomere is tightly defined
- deuterostome
- blastopore develops into the anus, with the second opening developing into the mouth
- diploblast
- animal that develops from two germ layers
- enterocoely
- mesoderm of deuterostomes develops as pouches that are pinched off from endodermal tissue, cavity contained within the pouches becomes coelom
- eucoelomate
- animal with a body cavity completely lined with mesodermal tissue
- indeterminate cleavage
- cleavage pattern in which individual blastomeres have the character of “stem cells,” and are not yet predetermined to develop into specific cell types
- protostome
- blastopore develops into the mouth of protostomes, with the second opening developing into the anus
- pseudocoelomate
- animal with a body cavity located between the mesoderm and endoderm
- radial cleavage
- cleavage axes are parallel or perpendicular to the polar axis, resulting in the alignment of cells between the two poles
- radial symmetry
- type of symmetry with multiple planes of symmetry, with body parts (rays) arranged around a central disk
- schizocoely
- during development of protostomes, a solid mass of mesoderm splits apart and forms the hollow opening of the coelom
- spiral cleavage
- cells of one pole of the embryo are rotated or misaligned with respect to the cells of the opposite pole
- triploblast
- animal that develops from three germ layers
